Impacto de los estudios moleculares en la ficología comparada mexicana: el caso de las macroalgas marinas
Impact of molecular studies on the mexican comparative phycology: marine macroalgae case
Jhoana Díaz-Larrea1*, Francisco F. Pedroche2 y Abel Sentíes1
1Área Ficología Comparada, Departamento de Hidrobiología, Universidad Autónoma Metropolitana-Iztapalapa.
2 Departamento de Ciencias Ambientales, Universidad Autónoma Metropolitana - Lerma.
*Correspondencia: jhoanadiazl@gmail.com
RESUMEN.
Se presenta un recuento de los avances en el conocimiento molecular de las especies de macroalgas marinas registradas para México. A la fecha se han realizado en el país 45 trabajos y la mayor parte de las contribuciones se han dedicado al grupo de las algas rojas, seguido de las algas pardas. La taxonomía y la filogenia son las principales áreas en la que estos trabajos se han enfocado, e incluyen la propuesta de nuevos arreglos taxonómicos, nuevos registros, nuevas especies para la ciencia y conespecificidad de especies. El mayor número de estudios, realizados en un intervalo de 19 años, se ha concentrado en el Pacífico mexicano (Baja California), seguido del Atlántico (Caribe mexicano). La mayoría de los estudios han procesado material recolectado en los estados de Baja California y Quintana Roo. Del estado de Tabasco no se han registrado trabajos hasta la fecha. Se ha utilizado mayormente la secuenciación de ADN como método, sólo tres trabajos utilizaron citogenética y uno de ellos los marcadores RAPDs. Los resultados obtenidos en los trabajos se han publicado fundamentalmente en revistas especializadas internacionales y en menor cuantía en tesis de licenciatura y posgrado.
Palabras clave: estudios moleculares, filogenia, macroalgas marinas, México, taxonomía.
ABSTRACT.
We analyze the progress of molecular knowledge in marine macroalgae from Mexico. There are, at this time, 45 published studies focused on red algae, followed by brown algae. The studies involved mainly taxonomy and phylogeny, and included proposals of new taxonomic arrangement, new records, new species and species con-specificity. The majority of the studies, carry out in 19 years, were done in Mexican Pacific (Baja California) followed by those in the Atlantic region (Mexican Caribbean). Processed material was collected primarily from Baja California and Quintana Roo states. There are no records from Tabasco state. DNA sequencing constitutes the most used method; only three papers used cytogenetics and one RAPDs. These results have been published mainly in specialized international journals and few of them as undergraduate or graduate thesis.
Key words: marine macroalgae, Mexico, molecular studies, phylogeny, taxonomy.
Introdución.
La “biodiversidad” se refiere a todas las formas en que la vida se manifiesta en la Tierra. En su sentido más amplio, la biodiversidad no se limita al número de especies que han existido en la historia de la vida, sino que también incluye desde la variación genética en individuos y poblaciones, hasta la diversidad de ecosistemas y biomas (Martínez-Meyer et al. 2014). Uno de los ecosistemas más diversos es el marino, sobre todo en lo que se refiere a la franja costera. México, considerado como el duodécimo país a nivel mundial con mayor superficie marina y una enorme riqueza ecosistémica y específica, posee un conocimiento muy limitado sobre el número, abundancia y distribución de las especies presentes; con excepción quizá de aquellas de interés comercial. Por lo general, la biodiversidad costera es mejor conocida que la oceánica o la de la plataforma continental (Hendrickx 1995).
El desarrollo de la biología molecular ha tenido, aproximadamente desde los años setenta, injerencia en los estudios de biología comparada (entendida ésta como la aproximación de la forma en espacio y tiempo), sin embargo, en los últimos 20 años se ha hecho evidente la aproximación de métodos moleculares y sus interpretaciones en los resultados de los trabajos de sistemática, biogeografía y de evolución (filogenia) en la biodiversidad terrestre y acuática de nuestro planeta. Por supuesto la ficología marina no es la excepción y en las siguientes líneas se expresa el impacto y un punto de vista sobre el impacto o consecuencias que ha tenido esta aproximación en la sistemática de las macroalgas marinas presentes en las costas mexicanas.
Primeramente, habría que diferenciar dos tareas importantes de la sistemática y taxonomía de las algas: La primera, es la urgente necesidad de concluir un inventario realista y confiable de las especies que habitan nuestro país. Por otra parte, está la tarea de formación de sistématas de manera integral y con un conocimiento profundo de los grupos que estudian. Las dos tareas están lejos de alcanzarse en países como México, en donde los recursos económicos se encuentran en otros lugares menos en las universidades e institutos de investigación no orientados al estudio del humano. El presente escrito se encuentra en el ámbito de la primera tarea y representa un intento por establecer una línea base en el conocimiento molecular de nuestras especies algales marinas. Esta línea base puede ser el punto de partida y comparación para analizar, en los años futuros, hacia dónde vamos y qué tan rápido hemos avanzado en la Taxonomía Molecular Alfa Asistida (MAAT, por sus siglas en inglés) (Cianciola et al. 2010).
Un sistema taxonómico basado en ADN ha sido sugerido desde hace ya varios años (Tautz et al. 2003) sin demérito de los sistemas respaldados por la morfología. Incluso desde una perspectiva integral, muchas fuentes de información se conjugan para determinar la identidad de un organismo en lo que denominamos especie. Si somos consecuentes con el pensamiento darwiniano, estas entidades deben ser el resultado de un análisis de ascendencia - descendencia mediante métodos filogenéticos robustos que identifiquen grupos monofiléticos.
Sin embargo, en el caso de México, la diversidad algal conocida ha sido el resultado de varias décadas de estudios fundados en una aproximación morfológica. Con esta visión se ha construido un primer panorama de la biodiversidad algal en nuestros litorales (Dreckmann 1998; Ortega et al. 2001; Pedroche et al. 2005, 2008). Con esa misma información también se ha tratado de hacer una aproximación a la distribución de grupos por regiones geográficas y a delimitar estas regiones, con base en la presencia o ausencia de algunas especies, géneros o familias, del porcentaje de las divisiones y su distribución por estados en la República mexicana (Pedroche et al. 1993; Pedroche y Sentíes 2003).
A partir de la introducción de una metodología molecular más accesible se han desarrollado trabajos que profundizan o complementan estas visiones morfológicas con datos de regiones particulares del ADN proveniente de algunos grupos selectos, como veremos más adelante. De esta manera se inicia una etapa en la que un sistema de taxonomía molecular se introduce en el pensamiento de la ficología mexicana, y en la que como complemento a las conocidas Unidades Taxonómicas Operativas (OTUs) se considera ahora el concepto de Unidades Taxonómicas Moleculares Operativas (MOTUs) (Blaxter y Floyd 2003). En ella, los especímenes se asignan, a través de una identidad molecular (secuencias particulares), a una “especie molecular”.
Este recuento de los avances en el conocimiento molecular de las especies registradas para México, no se hace con la finalidad de sustituir lo hasta ahora generado sino más bien en la búsqueda, en un futuro cercano, de una congruencia entre los nombres de las OTU estimadas por diversos métodos, con sus identidades moleculares (MOTU).
El marco molecular. Los marcadores moleculares son características detectables, directa o indirectamente (ya sea del ADN o de una proteína, respectivamente), en los individuos o poblaciones que indican la constitución parcial o total de un fragmento de ADN anónimo o de un gen en particular. Se han empleado para definir especies en grupos taxonómicos en los que la información proporcionada por la morfología es ambigua, pero también en la reconstrucción de filogenias, en filogeografía, en la evaluación de los flujos genéticos y en la estructura espacial de las poblaciones (García de León 2001; Oliveira 2001), pues ofrecen la posibilidad de disponer de una fuente ilimitada de caracteres puntuales y objetivos, además de poseer un carácter universal (Rentería Alcántara 2007). Precisamente es la variación o el polimorfismo de los marcadores moleculares lo que se usa en los estudios de diversidad genética. Dicha información puede ser utilizada para la caracterización de un organismo o un grupo de organismos de diferentes niveles taxonómicos (Avise 1994).
La información genética puede ser obtenida directa o indirectamente por diferentes técnicas, incluidas las inmunológicas, citogenéticas, electroforesis, secuenciación de proteínas, hibridación de ácidos nucleicos, la restricción, la amplificación y la secuenciación de porciones de ADN. Estas técnicas, desarrolladas en las últimas cuatro décadas, han permitido analizar de forma rápida y objetiva la variabilidad genética de un gran número de organismos (Amos y Hoelzel 1992; Olsen 1990). El empleo de secuencias moleculares, sin embargo, ha sido el método más ampliamente utilizado debido a la facilidad y rapidez en su obtención con distintos cebadores, a la reducción de costos, a su precisión y al hecho de que son objeto de una menor subjetividad en la interpretación de resultados (Díaz-Larrea et al. 2015). Otro aspecto relevante es la disponibilidad de la información en bases de datos especializadas (NCBI, BOLD) que permiten comparar a nivel universal los datos obtenidos.
En el estudio de las macroalgas marinas diversos marcadores moleculares han sido utilizados de manera exitosa en numerosos estudios de filogeografía, taxonomía, genética de la conservación, uso y aprovechamiento de recursos, filogenia, ecología, biogeografía y más recientemente genética de poblaciones (Ashley y Dow 1994; Leliaert et al. 2014), los que han permitido resolver diferentes problemas detectados a diferentes niveles jerárquicos, como la formación de complejos taxonómicos interespecíficos (especies crípticas) por la sobreposición de caracteres morfológicos, ya que muchos de los grupos presentan una alta o baja variabilidad morfológica que dificulta separar a las especies. Por no mencionar el uso diferenciado y subjetivo de los criterios morfológicos que deben aplicarse en la determinación de ciertos grupos en conflicto o la asignación de epítetos erróneos y la consecuente subestimación o sobre-estimación de la biodiversidad, que ha dejado una sistemática inestable en varios grupos de algas (Cianciola et al. 2010).
Los marcadores más frecuentemente utilizados a nivel mundial han sido los genes mitocondriales: cox1, cox2, cox3, espaciador cox2-cox3 (Destombe et al. 2010; García-Rodríguez et al. 2013; Pareek et al. 2010; Rueness 2005; Saunders 2005; Yang et al. 2008; Yow et al. 2011); los genes del cloroplasto: rbcL, psbA y el espaciador de la RuBisCO (Byrne et al. 2002; Destombe y Douglas 1991; Guillemin et al. 2008; Gurgel y Fredericq 2004; Gurgel et al. 2004; Iyer et al. 2005; Kim et al. 2006; Rueness 2005) y los genes nucleares: ITS1, ITS2, LSU, SSU (Bellorin et al. 2004; Goff et al. 1994)
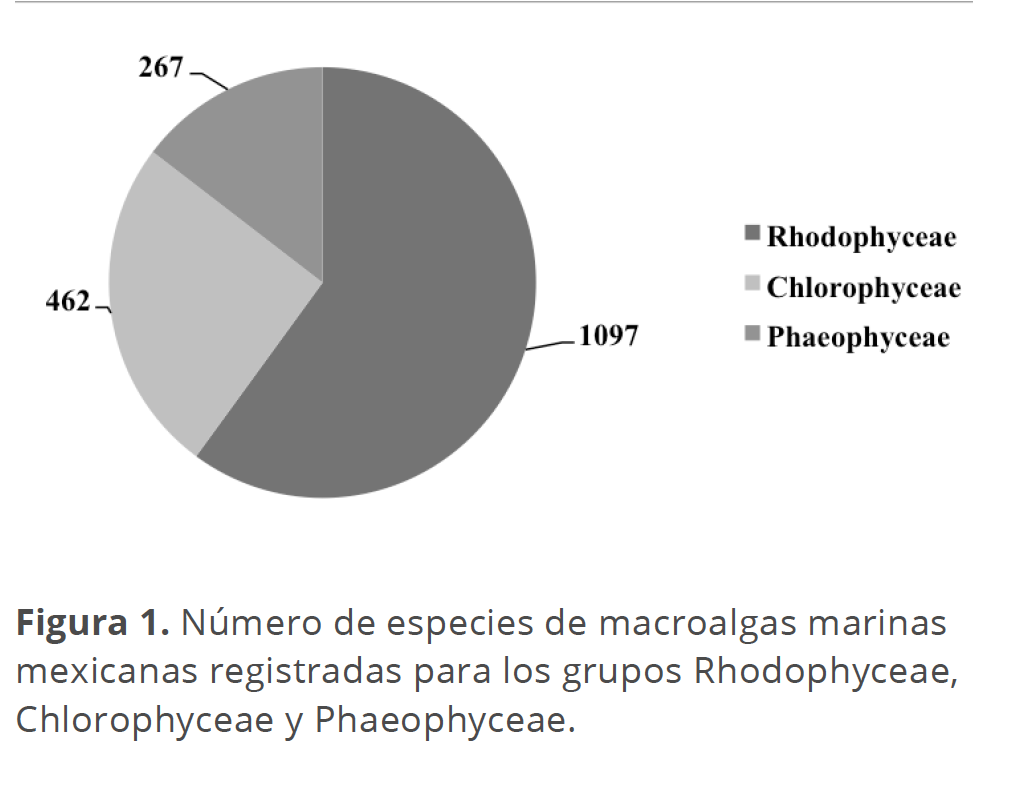
El efecto de la información molecular en la estimación de la biodiversidad algal marina de México. En nuestro país, la biodiversidad de macroalgas registrada al presente en los océanos Pacífico y Atlántico, comprende aproximadamente 1826 especies (Fig. 1): 1097 algas rojas (Rhodophyceae), 462 algas verdes (Chlorophyceae) y 267 algas pardas (Phaeophyceae) (Dreckmann 1998, Ortega et al. 2001; Pedroche y Sentíes 2003; Pedroche et al. 2005, 2008). Estos números comprenden fundamentalmente una aproximación basada en criterios morfológicos. Los trabajos en los que se ha abordado la diversidad biológica empleando marcadores moleculares son considerablemente escasos.
En el presente trabajo se presenta un análisis y revisión, casi exhaustiva, de los estudios de índole molecular que han empleado macroalgas marinas procedentes de los litorales mexicanos para la resolución de problemáticas diversas. Esto con el fin de ofrecer un panorama de las contribuciones de este enfoque nuevo a la comprensión de la biodiversidad algal y al establecimiento de líneas de investigación que aún están por ser exploradas.
Para lograr el objetivo planteado se realizó una búsqueda de artículos especializados y tesis publicadas desde 1997 a la fecha, en los que se utiliza información molecular para la solución de preguntas diversas en torno a la estimación de la diversidad de las macroalgas marinas que habitan las costas de México. Los autores se apoyaron en la base electrónica “Web of Science” para obtener un listado de artículos relacionados con el tema. Se completó la búsqueda en marzo de 2016, por lo que se incluyeron en las estadísticas del presente trabajo los registros encontrados hasta la fecha mencionada.
Así, al momento se han realizado en México un total de 45 trabajos (Cuadro 1), los cuales basan sus interpretaciones a partir de marcadores moleculares. La mayor parte de las contribuciones se han realizado en el grupo de algas rojas (58%) seguido de algas pardas (33%) y el menos abordado ha sido el grupo de las verdes (7%) (Fig. 2). Como se puede constatar, el número total de trabajos realizados es muy bajo si lo comparamos con el número total (1827) de macroalgas registradas al presente en los océanos Pacífico y Atlántico. Estos datos remarcan la importancia de comenzar a orientar los estudios en la ficología mexicana, bajo un enfoque integral de la sistemática, biogeografía y ecología con la biología molecular. Los grupos más abordados en estos trabajos de algas pardas son Sargassum, Padina y Dictyota, de las algas rojas son Gelidium, Bostrychia, complejo Laurencia, Porphyra y Pyropia y de las algas verdes el género Codium.
Las principales áreas en las que estos trabajos han tenido un impacto considerable han sido principalmente la taxonomía y filogenia, los cuales incluyen la propuesta de nuevos arreglos taxonómicos (Broom et al. 2002; Díaz-Larrea et al. 2007; León-Álvarez et al. 2014 a,b; Sentíes et al. 2015; Silva et al. 2004), nuevos registros (Aguilar-Rosas, L. et al. 2007, 2012; Aguilar-Rosas, R. et al. 2006, 2008; Lozano Orozco et al. 2014; Pedroche 1998; Sentíes et al. 2011; West et al. 2010; Zuccarello et al. 2012), conespecificidad de especies (Cassano et al. 2009), complejos de especies con enfoque filogeográfico (Núñez Reséndiz 2015; Núñez-Reséndiz et al. 2015) y nuevas especies para la ciencia (Lozano Orozco et al. 2015, 2016; Mateo-Cid et al. 2005, 2012, 2014, 2016; Mendoza-González et al. 2011; Reyes-Gómez 2015; Sentíes et al. 2014, 2015). Los enfoques de genética de poblaciones, filogeografía, ecología y biogeografía han sido menos abordados, probablemente por el sesgo que existe en la orientación de los ficólogos mexicanos, que es taxonómico. El alto costo de los estudios de genética de poblaciones y filogeografía ha limitado enormemente su uso en el campo de la ficología mexicana, aunado al limitado bagaje teórico previo existente (conocimiento de la biogeografía, filogenia molecular y taxonomía) de los grupos algales en cuestión. Sin embargo, los escasos trabajos realizados en biogeografía y ecología han permitido detectar especies introducidas (García-Rodríguez et al. 2013; Uwai et al. 2006), especies invasoras (Riosmena-Rodríguez et al. 2012) y patrones de distribución geográfica (López-Vivaz et al. 2015). En genética de poblaciones y filogeografía ha sido posible estimar la diversidad genética, el flujo genético y la estructura en poblaciones algales (García-Rodríguez et al. 2013; Hernández-Kantún 2014; Núñez-Reséndiz et al. 2015).
El mayor número de estudios (Fig. 3) se ha concentrado en el Pacífico mexicano, particularmente en Baja California, seguido del Atlántico en su porción del Caribe mexicano, en un intervalo de tiempo de 19 años (Fig. 4) que comprende desde el año 1997 hasta el 2016. En dicha figura se observa una clara tendencia hacia el incremento de estudios moleculares a partir del año 2011.
La mayoría de los estudios han abordado material recolectado en los estados de Baja California y Quintana Roo (Fig. 5), y los estados que presentan escasos trabajos (sólo 1) son Tamaulipas y Sonora. Del estado de Tabasco no se han registrado estudios hasta la fecha, lo que evidencia la necesidad de incrementar el conocimiento molecular en los 17 estados que presentan línea de costa en la República mexicana. Vale la pena destacar que incluso en aquellos estados donde se han realizado estudios, los sitios de recolecta visitados son escasos y no están distribuidos de manera homogénea.
Los géneros más abordados en estos trabajos son: Sargassum, Padina y Dictyota entre las Phaeophyceae, Gelidium, Bostrychia, el complejo Laurencia, Porphyra y Pyropia en las Rhodophyceae y entre de las Ulvophyceae hasta ahora sólo el género Codium.
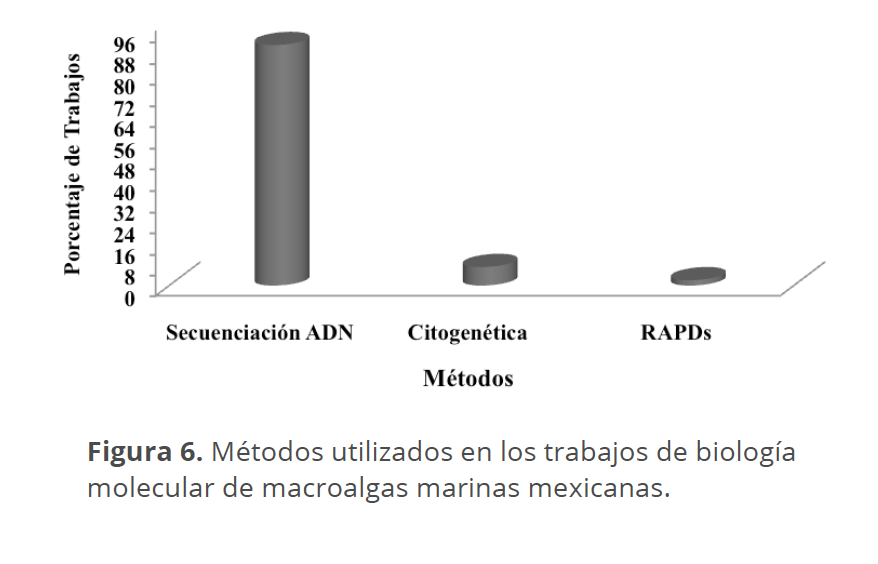
La mayor parte de los trabajos (Fig. 6) han utilizado la secuenciación de ADN, fundamentalmente plastidial y mitocondrial (Cuadro 1), debido al enfoque mayormente taxonómico y filogenético de los estudios. Sólo tres trabajos utilizaron citogenética (Ponce-Márquez et al. 1997, 2009), y uno de ellos los marcadores RAPDs (Ponce-Márquez 2010). La inclusión de otros métodos como los microsatélites permitiría abordar problemáticas diferentes a la taxonómica y filogenética.
La publicación de estos resultados se ha efectuado mayormente en revistas especializadas internacionales (80%) como Botanica Marina, y en menor cuantía en tesis, tanto de licenciatura como de posgrado (20%) (Fig. 7).
Finalmente, en cuanto a la participación institucional de las investigaciones, se puede decir que el trabajo de índole molecular se inicia en la UAM-Iztapalapa (Pedroche, 1998) y se ha continuado de manera colaborativa, con diversos laboratorios nacionales e internacionales. La UAM ha participado en el 51% del total de las publicaciones, la UABC en 22%, el IPN en 15%, la UNAM en 17%, la UABCS en 11% y la Universidad Michoacana de San Nicolás de Hidalgo en 6% (Fig. 8). Aunque estos números reflejan la presencia de ficólogos en estas instituciones, también habla de la infraestructura que diferencialmente caracteriza a nuestras instituciones de educación superior, resultado de políticas diversas y no coordinadas.
Consideraciones Finales.
En el presente trabajo queda evidenciado el impacto significativo pero escaso que han tenido los estudios moleculares en la ficología comparada mexicana. Una de las líneas potenciales de desarrollo es la participación en el proyecto Código de Barras, de la cual existen a nivel mundial varios grupos que abordan el “Barcoding” de algas, pero en México actualmente no existe ninguna red o grupo que aborde las algas con este enfoque. El enriquecimiento de otras líneas como la filogeografía y genética de poblaciones permitirá abordar procesos de especiación, eventos de radiación de especies, patrones biogeográficos, evaluar procesos de introducción y rastrear la ruta de dispersión de especies invasoras. Estos datos muestran la importancia de incentivar la naturaleza molecular de los estudios en la ficología mexicana, bajo un enfoque integral con la Sistemática, la Biogeografía y la Ecología. Sin embargo, esto sólo se puede alcanzar mediante la suma de voluntades, el trabajo colaborativo y la participación de instituciones diversas. Máxime cuando se trata de toda la flora algal de un país megadiverso, pues en esta contribución sólo se ha tocado lo referente a las algas marinas y en particular a las macroalgas.
La estimación de la biodiversidad debe ser una empresa en la que se suman los esfuerzos de los morfólogos, los taxónomos, los sistématas y los biólogos moleculares. Las instituciones, las asociaciones científicas, los gobiernos y la iniciativa privada, tienen un gran compromiso en el respaldo, el financiamiento y en la promoción de esta gran tarea.
REFERENCIAS.
Aguilar-Rosas, L.E., R. Aguilar-Rosas, H. Kawai, S. Uwai & E. Valenzuela-Espinoza. 2007. New record of Sargassum filicinum Harvey (Fucales, Phaeophyceae) in the Pacific Coast of Mexico. Algae 22: 17-21.
Aguilar-Rosas, L.E., S. Min Boo, K.M. Kim & C.V. Aguilar-Rosas. 2012. Primer registro de la especie japonesa Grateloupia turuturu (Halymeniaceae, Rhodophyta) en la costa del Pacífico mexicano. Hidrobiológica 22: 189-194.
Aguilar-Rosas, R., L.E. Aguilar-Rosas, G.Y. Cho & S.M. Boo. 2006. First record of Scytosiphon gracilis Kogame (Scylosiphonaceae, Phaeophyceae) for the Pacific coast of Mexico. Algae 21: 11-13.
Aguilar-Rosas, R., L.E. Aguilar-Rosas & S. Shimada. 2008. First record of Ulva pertusa Kjellman (Ulvales, Chlorophyta) in the pacific coast of Mexico. Algae 23: 201-207.
Amos, B. & A.R. Hoelzel. 1992. Applications of molecular genetic techniques to the conservation of small populations. Biology Conservation 61: 133-144.
Ashley, M.V. & B.D. Dow. 1994. The use of microsatellite analysis in population biology: background, methods and potential applications. In: B. Schierwater, B. Streit, G.P. Wagner & R. DeSalle. Eds. Molecular Ecology and Evolution: Approaches and Applications. Birkhauser Verlag, Basel, pp. 185-201.
Avise, J.C. 1994. Molecular markers, natural history and evolution. Chapman & Hall, Inc., New York.
Bellorin, A.M., M.C. Oliveira & E.C. Oliveira. 2004. Gracilaria vermiculophylla: a western Pacific species of Gracilariaceae (Rhodophyta) first recorded from the eastern Pacific. Phycological Research 52: 69-79.
Blaxter, M. & R. Floyd. 2003. Molecular taxonomics for biodiversity surveys: already a reality. Trends in Ecology & Evolution 18: 268-269.
Broom, J.E., W.A. Nelson, C. Yarish, W.A. Jones, R. Aguilar Rosas & L.E. Aguilar-Rosas. 2002. A reassessment of the taxonomic status of Porphyra suborbiculata, Porphyra carolinensis and Porphyra lilliputiana (Bangiales, Rhodophyta) based on molecular and morphological data. European Journal of Phycology 37: 227-235.
Byrne, K., G.C. Zuccarello, J. West, M.L. Liao. & G.T. Kraft. 2002. Gracilaria species (Gracilariaceae, Rhodophyta) from southeastern Australia, including a new species, Gracilaria perplexa sp. nov.: morphology, molecular relationships and agar content. Phycological Research 50: 295-311.
Cassano, V., J. Díaz-Larrea, A. Sentíes, M.C. Oliveira, M.C. Gil-Rodríguez & M.T. Fujii. 2009. Evidence for the conspecifity of Palisada papillosa with P. perforata (Ceramiales, Rhodophyta) from the western and eastern Atlantic Ocean on the basis of morphological and molecular analyses. Phycologia 48: 86-100.
Cianciola, E., T. Popolizio, C. Schneider. & C. Lane. 2010. Using molecular-assisted alpha taxonomy to better understand red algal biodiversity in Bermuda. Diversity 2: 946-958.
Destombe, C. & S.E. Douglas. 1991. RuBisCO spacer sequence divergence in the rhodophyte alga Gracilaria verrucosa and closely related species. Current genetics 19: 395-8.
Destombe, C., M. Valero & M.L. Guillemin. 2010. Delineation of two sibling red algal species, Gracilaria gracilis and Gracilaria dura (Gracilariales, Rhodophyta), using multiple DNA markers: resurrection of the species G. dura previously described in the Northern Atlantic 200 years ago. Journal of Phycology 46: 720-727.
Díaz-Larrea, J. 2008. Sistemática molecular del complejo Laurencia (Ceramiales, Rhodophyta) en el Caribe mexicano. Tesis de Doctorado. Universidad Autónoma Metropolitana, Iztapalapa. 131 pp.
Díaz-Larrea, J., A. Sentíes, M.T. Fujii, F.F. Pedroche & M.C. Oliveira. 2007. Molecular evidence for Chondrophycus poiteaui var. gemmiferus comb. et stat. nov. (Ceramiales, Rhodophyta) from the Mexican Caribbean Sea: implications for the taxonomy of the Laurencia complex. Botanica Marina 50: 250-256.
Díaz-Larrea, J., A. Sentíes, M.L. Núñez-Reséndiz, M.L. López-Valdez. & K.M. Dreckmann. 2015. La genética de poblaciones como herramienta útil en diversos estudios de macroalgas marinas. Cymbella 1: 1-11.
Díaz-Martínez, S. 2008. Estudio morfométrico y genético-molecular de Padina gymnospora (Kützing) Sonder. Tesis de Licenciatura. Facultad de Estudios Superiores Zaragoza, Universidad Nacional Autónoma de México. 56 pp.
Díaz-Martínez, S. 2011. Evaluación taxonómica de las morfoespecies de Padina (Dictyotaceae) en el Atlántico y Pacífico Tropical de México con base en datos moleculares. Tesis de Maestría. Facultad de Estudios Superiores Zaragoza, Universidad Nacional Autónoma de México. 94 pp.
Dreckmann, K.M. 1998. Clasificación y nomenclatura de las macroalgas marinas bentónicas del Atlántico mexicano. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO), México.
Fernández García, C. 2012. Taxonomía y biogeografía de las familias Caulerpaceae (Chlorophyta), Dictyotaceae (Ochrophyta) y Corallineaceae (Rhodophyta) en el Pacífico de Centroamérica. Tesis de Doctorado. Universidad Autónoma de Baja California Sur. 271 pp.
García-Rodríguez, L.D., R. Riosmena-Rodríguez, S.Y. Kim, M. López-Meyer, J. Orduña-Rojas, J.M. López-Vivas & S.M. Boo. 2013. Recent introduction of Gracilaria parvispora (Gracilariales, Rhodophyta) in Baja California, Mexico. Botanica Marina 56: 143-150.
García de León, F.J. 2001. Los marcadores genéticos en el conocimiento y manejo de recursos bióticos. Biotam 12: 57-80.
Goff, L.J., D.A. Moon. & A.W. Coleman. 1994. Molecular delineation of species and species relationships in the red algal agarophytes Gracilariopsis and Gracilaria (Gracilariales). Journal of Phycology 30: 521-37.
Guillemin, M.L., S.A. Akki, T. Givernaud, A. Mouradi, M. Valero & C. Destombe. 2008. Molecular characterization and development of rapid molecular methods to identify species of Gracilariaceae from the Atlantic coast of Morocco. Aquatic Botany 89: 324-30.
Gurgel, C.F.D. & S. Fredericq. 2004. Systematics of the Gracilariaceae (Gracilariales, Rhodophyta): A critical assessment based on rbcL sequence analyses. Journal of Phycology 40: 138-159.
Gurgel, C.F.D., S. Fredericq & J.N. Norris. 2004. Phylogeography of Gracilaria tikvahiae (Gracilariaceae, Rhodophyta): a study of genetic discontinuity in a continuously distributed species based on molecular evidence. Journal of Phycology 40: 748-758.
Hammann, M., G. Wang, S.M. Boo, L.E. Aguilar‑Rosas & F. Weinberger. 2016. Selection of heat‑shock resistance traits during the invasion of the seaweed Gracilaria vermiculophylla. Marine Biology 163: 104.
Hendrickx, M.E. 1995. Camarones. In: W. Fischer, F. Krupp, W. Schneider, C. Sommer, K.E. Carpenter & V.H. Niem. Eds. Guía FAO para la identificación de especies para los fines de la pesca. Pacífico centro-oriental. Vol. 1. Plantas e invertebrados. FAO, Roma, pp. 417-537.
Hernández-Kantún, J.J., R. Riosmena-Rodriguez, W.H. Adey & F. Rindi. 2014. Analysis of the cox2-3 spacer region for population diversity and taxonomic implications in rhodolith-forming species (Rhodophyta: Corallinales). Phytotaxa 190: 331-354.
Iyer, R., E.M. Tronchin, J.J. Bolton & V.E. Coyne. 2005. Molecular systematics of the Gracilariaceae (Gracilariales, Rhodophyta) with emphasis on Southern Africa. Journal of Phycology 41: 672-684.
Kim, M.S., E.C. Yang & S.M. Boo. 2006. Taxonomy and phylogeny of flattened species of Gracilaria (Gracilariaceae, Rhodophyta) from Korea based on morphology and protein-coding plastid rbcL and psbA sequences. Phycologia 45: 520-528.
Leliaert, F., H. Verbruggen, P. Vanormelingen, F. Steen, J.M. López-Bautista, G.C. Zuccarello & O. De Clerck. DNA-based species delimitation in algae. 2014. European Journal of Phycology 49: 179-196.
León-Alvarez, D., M.L. Núñez-Reséndiz & M.E. Ponce-Márquez. 2014a. Morphological and molecular characterization of Neoralfsia hancockii comb. nov. (Ralfsiales, Phaeophyceae) from topotype of San José del Cabo, Baja California, Mexico. Botanica Marina 57: 139-146.
León-Alvarez, D., M.L. Núñez-Reséndiz & M.J. Wynne. 2014b. Morphological and molecular studies on topotype material of Neoralfsia expansa (Phaeophyceae) reveal that Asian specimens assigned to this taxon are genetically distinct. Botanica Marina 57: 351-358.
Lindstrom, S.C., J.R. Hughey & L.E. Aguilar Rosas. 2015. Four new species of Pyropia (Bangiales, Rhodophyta) from the west coast of North America: the Pyropia lanceolata species complex updated. PhytoKeys 52: 1-22.
Lopez-Vivas, J.M., R. Muñiz-Salazar, R. Riosmena-Rodriguez, I. Pacheco-Ruiz & C. Yarish. 2015. Endemic Pyropia species (Bangiales, Rhodophyta) from the Gulf of California, Mexico. Journal of Applied Phycology 27: 1029-1041.
Lozano Orozco, J.G. El género Dictyota (Phaeophyceae: Dictyotales) en el Atlántico mexicano: análisis morfológico y molecular. Tesis de Doctorado. Universidad Autónoma Metropolitana, Iztapalapa. 138 p.
Lozano Orozco, J.G., A. Sentíes, J. Díaz-Larrea, F.F. Pedroche & O. De Clerck. 2014. The occurrence of Dictyota canariensis (Dictyotales, Phaeophyceae) in the Gulf of Mexico. Botanica Marina 57: 359-365.
Lozano Orozco, J.G., A. Sentíes, O. De Clerck, K.M. Dreckmann & J. Díaz-Larrea. 2015. Two new species of the genus Dictyota (Phaeophyceae: Dictyotales) from the Mexican Caribbean. American Journal of Plant Sciences 6: 2492-2501.
Lozano-Orozco, J.G., A. Sentíes, J. Díaz-Larrea, & F.F. Pedroche. 2016. Dictyota chalchicueyecanensis sp. nov. (Dictyotales; Phaeophyceae) en el Golfo de México: Evidencias moleculares y morfológicas. Hidrobiológica No. especial SOFILAC 26: (en prensa).
Martínez-Meyer, E., J.E. Sosa-Escalante & F. Álvarez. 2014. El estudio de la biodiversidad en México: ¿una ruta con dirección? Revista Mexicana de Biodiversidad, Supl. 85: S1-S9.
Mateo-Cid, L.E., A.C. Mendoza-González, J. Díaz-Larrea, A. Sentíes, F.F. Pedroche & J.D. Sánchez Heredia. 2012. A new species of Pyropia (Rhodophyta, Bangiaceae), from the Pacific coast of Mexico, based on morphological and molecular evidence. Phytotaxa 54: 112.
Mateo-Cid, L.E., A.C. Mendoza-González, B. Gavio & S. Fredericq. 2005. Grateloupia huertana sp. nov. (Halymeniaceae, Rhodophyta) a peculiar new postrate species from tropical Pacific Mexico. 2005. Phycologia 44: 4-16.
Mateo-Cid, L.E., A.C. Mendoza-González, A. Sentíes, J. Díaz-Larrea & J.A. Acosta Calderón. 2014. Laurencia laurahuertana sp. nov. (Rhodomelaceae, Rhodophyta); an epiphytic species from the Mexican Caribbean. Phycological Research 62: 94-101.
Mateo-Cid, L.E. A.C. Mendoza-González, A. Sentíes, J. Díaz-Larrea, D.Y. García-López & B. Martínez-Daranas. 2016. Description of two new species of Ochtodes (Rhodophyta: Gigartinales) from Caribbean Mexico on the basis of morphological and molecular evidence. Botanica Marina 59 (en prensa).
Mendoza-González, C.A., A. Sentíes, L.E. Mateo-Cid, J. Díaz-Larrea, F.F. Pedroche & R. Alvarado Villanueva. 2011. Ochtodes searlesii sp. nov. (Gigartinales, Rhodophyta), from the Pacific tropical coast of Mexico, based on morphological and molecular evidence. Phycological Research 59: 250-258.
Núñez Reséndiz, M.L. 2015. Variación genética de las poblaciones de Hydropuntia cornea (Gracilariaceae, Rhodophyta) en las costas del Golfo de México y el Caribe mexicano. Tesis de Doctorado. Universidad Autónoma Metropolitana, Iztapalapa. 163 pp.
Núñez-Reséndiz, M.L., K.M. Dreckmann, A. Sentíes, J. Díaz-Larrea & G.M. Zuccarello. 2015. Genetically recognizable but not morphologically: The cryptic nature of Hydropuntia cornea and H. usneoides (Gracilariales, Rhodophyta) in the Yucatan Peninsula. Phycologia 54: 407-416.
Oliveira, M.C. 2001. Estudios de la macro y microbiodiversidad de las algas. Secuenciamiento del DNA ribosomal (rDNA). In: K. Alveal, & T. Antezana. Eds. Sustentabilidad de la biodiversidad. Bases científico-técnicas, teorizaciones y proyecciones. Ed. de la Universidad de Concepción, Concepción, pp. 85-96.
Olsen, J.L. 1990. Nucleic acids in algal systematics. Journal of Phycology 26: 209-214.
Ortega, M.M., J.L. Godínez & G. Garduño-Solórzano. 2001. Catálogos de algas bénticas de las costas mexicanas del Golfo de México y Mar Caribe. Universidad Nacional Autónoma de México y Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, México.
Pareek, M., A. Mishra & B. Jha. 2010. Molecular phylogeny of Gracilaria species inferred from molecular markers belonging to three different genomes. Journal of Phycology 46: 1322-1328.
Pedroche, F.F. 1998. El género Codium (Chlorophyta) en el Pacífico de México. Tesis de Doctorado. Facultad de Ciencias, Universidad Nacional Autónoma de México. 129 pp.
Pedroche, F.F. 2001. Estudios filogenéticos del género Codium (Chlorophyta) en el Pacífico mexicano. Uso de ADNr mitocondrial. Anales de la Escuela Nacional de Ciencias Biológicas 47(1): 109-123.
Pedroche F.F., K.M. Dreckmann, A. Sentíes & R. Margain-Hernández. 1993. Diversidad algal en México. Revista Mexicana de Historia Natural Vol. Esp. 44: 69-92.
Pedroche, F.F. & A. Sentíes. 2003. Ficología marina mexicana. Diversidad y Problemática actual. Hidrobiológica 13: 23-32.
Pedroche, F.F., P.C. Silva, L.E. Aguilar-Rosas, K.M. Dreckmann & R. Aguilar-Rosas. 2005. Catálogo de las Algas Marinas Bentónicas del Pacífico de México I. Chlorophycota. Universidad Autónoma Metropolitana, Universidad Autónoma de Baja California, University of California-Berkeley. Mexicali.
Pedroche, F.F. P.C. Silva, L.E. Aguilar-Rosas, K.M. Dreckmann, & R. Aguilar-Rosas. 2008. Catálogo de las algas bentónicas del Pacífico de México II. Phaeophycota, Universidad Autónoma Metropolitana, Universidad Autónoma de Baja California, University of California-Berkeley, Mexicali.
Ponce-Márquez, M.E. 2010. Gelidium (Gelidiales, Rhodophyta) del Pacífico tropical mexicano: variación genética inter e intra específica. Tesis de Doctorado. Universidad Nacional Autónoma de México. 142 pp.
Ponce Márquez, M.E., A. Santamaría, D. Rodríguez Vargas & A. Sarmiento Montiel. 1997. Estudio citogenético de macroalgas marinas con posibilidades de explotación. Oceanología 13: 155-166.
Ponce-Márquez, M.E., D. Rodríguez, N.M. López, L.A. Lois & M.A. Ángeles Aguilar. 2009. Estudio citogenético de poblaciones mexicanas de Gelidium sclerophyllum (Gelidiales, Rhodophyta). Hidrobiológica 19: 85-93.
Rentería Alcántara, M. 2007. Breve revisión de los marcadores moleculares. In. L.E. Eguiarte, V. Souza y X. Aguirre. Comps. Ecología molecular. Secretaría del Medio Ambiente y Recursos Naturales, Instituto Nacional de Ecología, Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. México, pp. 541-571.
Reyes-Gómez, V.P. 2015. Caracterización morfológica y molecular de las algas costrosas pardas gelatinosas del Pacífico Tropical Mexicano. Tesis de Doctorado. Universidad Nacional Autónoma de México. 106 pp.
Riosmena-Rodríguez, R., G. Hun Boo, J.M. López-Vivas, A. Hernández-Velasco, A. Sáenz-Arroyo & S. Ming Boo. 2012. The invasive seaweed Sargassum filicinum (Fucales, Phaeophyceae) is on the move along the Mexican Pacific coastline. Botanica Marina 55: 547-551.
Rueness, J. 2005. Life history and molecular sequences of Gracilaria vermicullophylla (Gracilariales, Rhodophyta), a new introduction to European waters. Phycologia 44: 120-128.
Saunders, G.W. 2005. Applying DNA barcoding to red macroalgae: a preliminary appraisal holds promise for future application. Philosophical Transactions of the Royal Society Series B 360: 1879-1888.
Sentíes, A., J. Díaz-Larrea, V. Cassano, M.C. Gil-Rodríguez & M.T. Fujii. 2011. Laurencia marilzae (Ceramiales, Rhodophyta) from the Mexican Caribbean: A new record for the tropical western atlantic. Bulletin of Marine Sciences 87: 681-686.
Sentíes, A., A.C. Mendoza-González, L.E. Mateo-Cid, J. Díaz-Larrea, G.A. Ceballos-Corona & M.T. Fujii. 2014. Osmundea purepecha sp. nov. (Rhodophyta: Ceramiales) from the Pacific tropical coast of Mexico, based on morphological and molecular data. Pacific Sciences 68: 295-304.
Sentíes, A., M.J. Wynne, V. Cassano, J. Díaz-Larrea, M.C. Gil-Rodriguez & M.T. Fujii. 2015. Yuzurua iridescens (M.J. Wynne & D.L. Ballantine) comb. nov. (Ceramiales, Rhodophyta) from the Caribbean Sea: morphological and molecular evidence. Brazilian Journal of Botany 38: 605-613.
Silva, P.C., F.F. Pedroche, M.E. Chacana, R. Aguilar-Rosas, L.E. Aguilar-Rosas & J. Raum. 2004. Geographic correlation of morphological and molecular variation in Silvetia compressa (Fucaceae, Fucales, Phaeophyceae). Phycologia 43: 204-214.
Tautz, D., P. Arctander, A. Minelli, R.H. Thomas & A.P Vogler. 2003. A plea for DNA taxonomy. Trends in Ecology & Evolution 18: 70-74.
Uwai, S., W. Nelson, K. Neil, W.D. Wang, L.E. Aguilar-Rosas, S.M. Boo, T. Kitayama & H. Kawai. 2006. Genetic diversity in Undaria pinnatifida (Laminariales, Phaeophyceae) deduced from mitochondria genes-origins and succession of introduced populations. Phycologia 45: 687-695.
Vázquez-Delfín, E., G.H. Boo, D. Rodríguez, S.M. Boo & D. Robledo. 2016. Hypnea musciformis (Cystocloniaceae) from the Yucatan Peninsula: morphological variability in relation to life-cycle phase. Phycologia 55: 230-242.
West, J.A., G.C. Zuccarello, F.F. Pedroche & S.L. de Goer. 2010. Rosenvingea orientalis (Scytosiphonaceae, Phaeophyceae) from Chiapas, Mexico: life history in culture and molecular phylogeny. Algae 25: 187-195.
Yang, E.C., M.S. Kim, P.J.L. Geraldino, D. Sahoo, J.A. Shin & S.M. Boo. 2008. Mitochondrial cox1 and plastid rbcL genes of Gracilaria vermiculophylla (Gracilariaceae, Rhodophyta). Journal of Applied Phycology 20: 161-168.
Yow, Y.Y., P.E. Lim. & S.M. Phang. 2011. Genetic diversity of Gracilaria changaii (Gracilariaceae, Rhodophyta) from west coast Peninsular Malaysia based on mitochondrial cox1 gene analysis. Journal of Applied Phycology 23: 219-226.
Zuccarello, G.C., J. Buchanan, J.A. West & F.F. Pedroche. 2011. Genetic diversity of the mangrove-associated alga Bostrychia radicans/Bostrychia moritziana (Ceramiales, Rhodophyta) from southern Central America. Phycological Research 59: 98-104.
Zuccarello, G.C., M. Kamiya, R. Ootsuki, S. Loiseaux de Goër, F.F. Pedroche & J.A. West. 2012. New records of red algae from mangroves in El Salvador and Pacific Mexico, combining culture and molecular observations. Botanica Marina 55: 101-111.
Recibido: 8 de abril
Revisado: 20 de mayo
Corregido: 9 de junio
Aceptado: 15 de junio
Revisores: Dos revisores anónimos.








 Vol. 2, No. 1
Vol. 2, No. 1