Las interpretaciones de los taxones, uno de los problemas de la taxonomía tradicional. Caso: Scenedesmus sensu lato (Sphaeropleales, Chlorophyceae).
Taxa interpretations, one of the problems of traditional taxonomy. Case: Scenedesmus sensu lato (Sphaeropleales, Chlorophyceae).
Augusto A. Comas González
Centro de Estudios Ambientales de Cienfuegos, Apartado Postal 5, C.P. 59350. Cienfuegos, Cuba.
E-mail:comasaugusto@gmail.com; augusto@gestion.ceac.cu<
Comas González. A.A. 2020. Las interpretaciones de los taxones, un de los problemas de la taxonomía tradicional. Caso: Scendesmus sensu lato (Sphaeropleales, Chlorophyceae). Cymbella 6(3): 139-150.
INTRODUCCIÓN.
La taxonomía tradicional presenta serias limitaciones, entre ellas, su incapacidad de dar respuesta a la amplia variabilidad de muchas especies, en consecuencia, se han descrito taxones con caracteres diagnósticos y límites imprecisos, tipos no designados o los seleccionados por autores posteriores, con frecuencia, no se corresponden exactamente con las descripciones originales, lo que ha propiciado diferentes interpretaciones complicando la taxonomía de los taxones involucrados, especialmente su nomenclatura.
Interpretación (acción y efecto de interpretar) posee varias acepciones, me adscribo a la de: “concebir, ordenar o expresar de un modo personal la realidad” que, en mayor o menor grado, está siempre presente en la taxonomía tradicional. Me referiré exclusivamente a aquellas interpretaciones sobre algunas especies de Scenedesmus sensu lato cuyos caracteres diagnósticos y límites específicos son imprecisos, originando diferentes criterios taxonómicos.
No obstante, creo conveniente mencionar brevemente dos conceptos clásicos en la taxonomía de cianobacterias y microalgas:1) F. Drouet (1907-1982), ficólogo norteamericano dedicado al estudio de las cianofíceas (cianobacterias), seguía el criterio de taxones muy amplios con una gran variabilidad morfológica, de ello, la existencia de pocos géneros que incluían un número reducido de especies y 2) T. Hortobagyi (1912-1990), ficólogo húngaro dedicado a las Chlorococcales sensu lato, quien, a diferencia del anterior autor, concebía taxones en un sentido muy estrecho, cada morfotipo constituía un taxón, ya sea especie, variedad o forma. Era fiel exponente de la llamada concepción politética (Komárek 1978).
Tampoco incluyo a los conceptos o principios de trabajo (no a los conceptos o definiciones de especies), a la hora de definir cuándo se trata de una especie, variedad o forma.
Hasta ahora, la nomenclatura de las algas ha sido regulada por el Código Internacional de Nomenclatura Botánica. Tanto en sus diferentes ediciones como en el actual Código Internacional de Nomenclatura de algas, hongos y plantas, se ha permitido que los holotipos de las especies de microalgas, el tipo nomenclatural, sea una ilustración, denominado iconotipo o “typus tabla”, aportando ventajas y desventajas. El material tipo conservado en ejemplares de herbarios, ya prácticamente en desuso, no era un ejemplar como en las plantas superiores, sino una mezcla de especímenes, no solo de una misma entidad, sino de diferentes. El iconotipo aclara cuál taxón es el que se describe, descartando al resto, sin embargo, antes de la invención de las fotografías, la “fidelidad” o representación exacta de la especie en cuestión, dependía de las habilidades del dibujante y por supuesto, de una selección adecuada, en correspondencia con la diagnosis. Ejemplo de inmejorables ilustraciones son las aportadas por los trabajos de Skuja y Hortobagyi. El iconotipo debía representar el espécimen más representativo, lo que, generalmente, no ha ocurrido. En la actualidad se designan cultivos tipo debidamente conservados y depositados en instituciones reconocidas.
Podemos afirmar que de las diferentes interpretaciones surgen de taxones con descripciones escuetas, con definiciones imprecisas de los caracteres diagnósticos y de los diacríticos, es decir, de aquellos por los cuales se diferencian de los taxones más relacionados. En muchos casos los tipos nomenclaturales no se han designado, se han extraviado, o no coinciden plenamente con las diagnosis.
Principales características y breve historia del género Scenedesmus.
Scenedesmus Meyen 1829, es uno de los géneros morfológicos dentro de las clorofíceas cocales más distribuido en las aguas dulces del orbe. Por su particular morfología, salvo excepciones, se reconoce fácilmente de los demás géneros: i) forma colonias especiales (cenobios) más o menos lineales, en una o dos hileras, o marcadamente alternadas, ii) formas celulares diversas, desde obtusas, cilíndricas, ovadas, ovales hasta fusiformes, iii) algunas especies con deposiciones en la pared celular o con estructuras específicas de esta, como espinas, dentículos, costillas, verrugas, etc. iv) con un cloroplastidio parietal con un pirenoide y v) presenta una reproducción por autosporas unidas en autocenobios, aunque existen algunos registros de formación de estadios flagelares (Lukavský 1991, Trainor 1963).
A pesar de su relativa facilidad para ser reconocido como género, muchas de sus especies, por su amplia variabilidad morfológica y, por ende, con una complicada taxonomía, han constituido un reto para los taxónomos y para los que precisan de una correcta identificación.
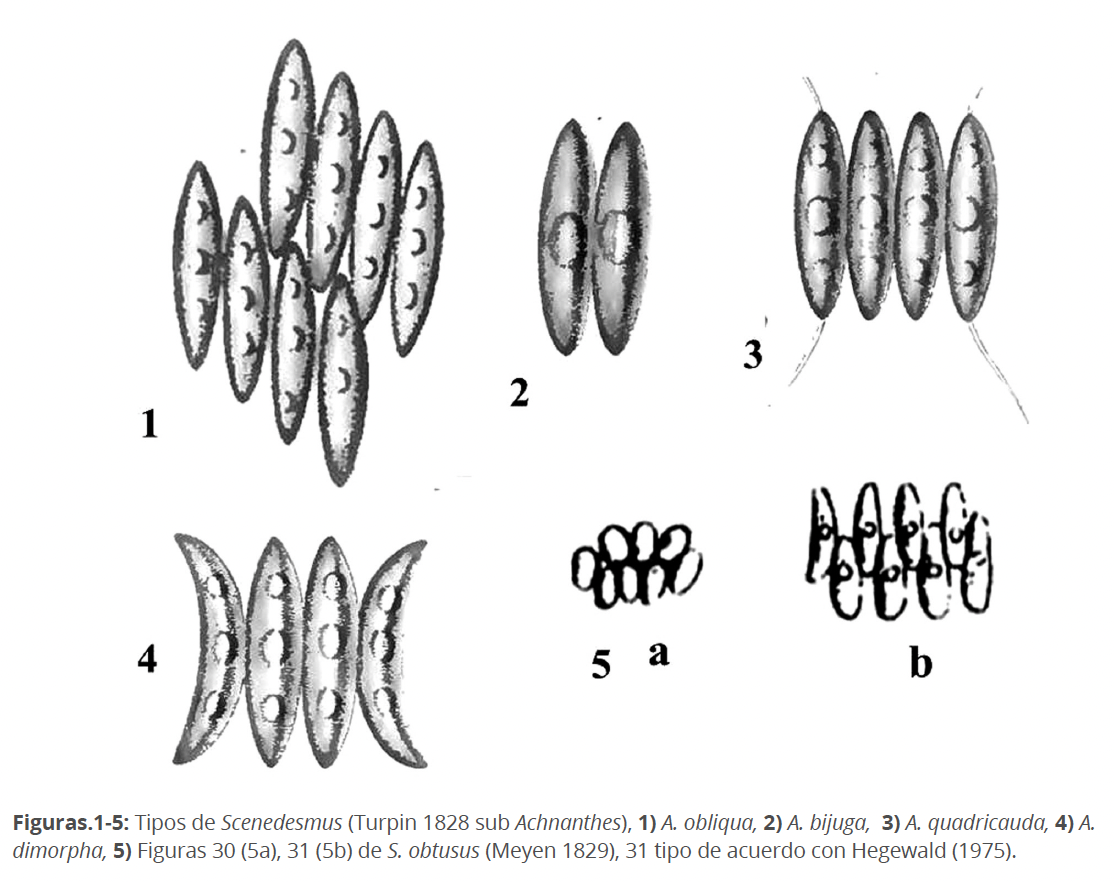
Pudiéramos afirmar que Scenedesmus tuvo un origen “sui generis”. Hasta ahora, las consideradas sus primeras especies, fueron descritas dentro de la diatomea Achnanthes (Turpin 1828). Sus ilustraciones eran muy esquemáticas, estilizadas, además las formas celulares eran muy similares entre ellas (Figs. 1-4). El autor para la identificación de las especies hizo énfasis en el número y disposición de las células en la colonia, así como en la presencia, al menos en una de ellas, de espinas en los polos de las células marginales. Véase que entre A. bijuga y A. obliqua (Figs. 1-2), la diferencia estriba en el número y disposición de las células. A. quadricauda se distingue por la presencia de espinas en los polos de las células marginales (Fig. 3) y A. dimorpha, por la curvatura de las células marginales (Fig. 4).
En la descripción original, Meyen (1829) no designa tipo. La etimología del nombre proviene del griego: σκηνή (skeni) = cinta, celda, tienda, δεσμός (desmos) = unión, ligadura). Su diagnosis fue simple: “Alga acintada, sin talo propio, formada por células elípticas iguales agregadas lateralmente en una seria plana. Masa interna regularmente dispuesta, de color verde, grumosa de márgenes doblados”. Su ortografía fue relevantemente controvertida: Meyen (1829), en el texto escribe Scenedesmus; pero en las láminas Scaenaedesmus. Ehrenberg (1834), propone, sobre bases idiomáticas, Scenodesmus. A pesar de esta fundamentada corrección, el nombre Scenedesmus (aplicado por Kützing en 1849, ha prevalecido hasta hoy. Nägeli (1949) designa a S. obtusus Meyen como tipo del género; pero lo escribe como Scenodesmus. El tipo seleccionado incluye las Figs. 30 y 31 de Meyen (1829), que actualmente representan, sin lugar a duda a dos especies diferentes. La Fig. 5a (Fig.30 de Meyen 1829), es muy similar a S. verrucosus Roll. La Fig. 5b (Fig.31 de Meyen 1829), fue seleccionada por Hegewald et al. (1975) como tipo del género.
Aunque el objetivo fundamental de este artículo es mostrar las diferentes interpretaciones con algunos ejemplos, creo necesario referirme a algunos aspectos generales. Las primeras aportaciones a la taxonomía se fundamentan en criterios morfológicos, luego se tomó en cuenta a la ultraestructura de la pared celular, así como a su ecología, distribución geográfica, características bioquímicas y más recientemente en los análisis de sus filogenias moleculares.
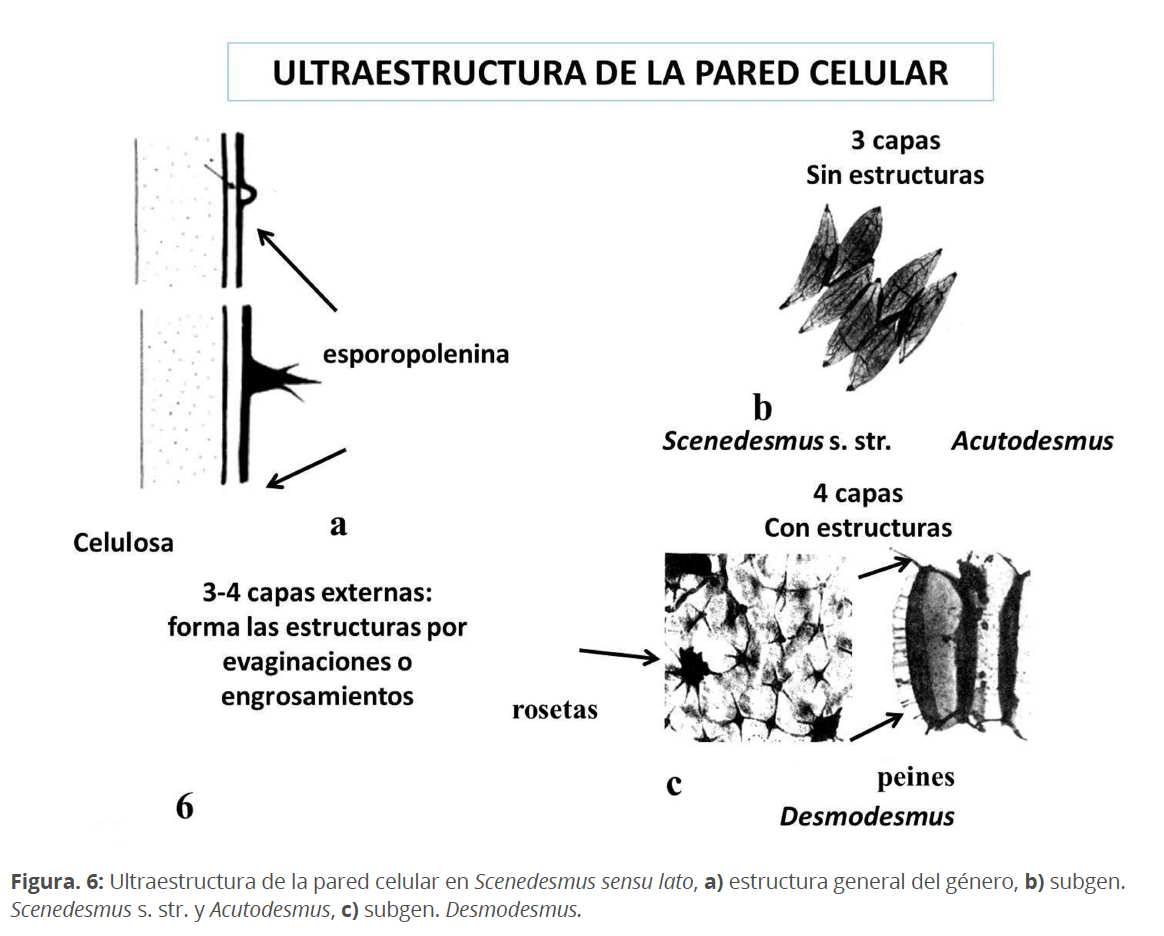
La taxonomía tradicional demostró que dentro del género era posible diferenciar varios grupos. De esta manera se propusieron diferentes sistemas dentro del taxón, algunos complicados, definiendo subgéneros, secciones, series, etc. Entre los más recientes tenemos a los sistemas de Hegewald & Schepf (1978) y Komárek & Fott (1983). En general diferencian tres grupos principales a nivel de subgéneros: sin estructuras de la pared como espinas, dientes, etc. Scenedesmus y Acutodesmus, este último caracterizado por células en principio fusiformes con sus extremos más o menos aguzados; dentro de Desmodesmus se incluyen las especies con estructuras parietales.
Los avances en la microscopía electrónica permitieron confirmar la composición de la pared celular: una capa interna de celulosa y externamente, 3-4 capas de esporopolenina. (Fig. 6). Los subgéneros Scenedesmus y Acutodesmus presentan 3 capas de esporopolenina sin ningún típico de estructuras. Si al microscopio fotónico se observaban formaciones semejantes a costillas o granulaciones, estas no eran verdaderas estructuras, sino deposiciones. Desmodesmus presenta, por otro lado, 4 capas, la más externa formando estructuras diversas. Se definieron no solo las espinas, dentículos y costillas, sino también otras estructuras no observadas al microscopio fotónico. De esta manera, se identificaron las denominadas rosetas (a veces observadas al microscopio fotónico como diminutas estructuras tubulares), así como ornamentaciones reticuladas, patrones, mediante los cuales pudieran atribuirse a determinadas especies, por tanto, de indiscutible valor taxonómico (Bisalputra & Weier 1963, Hegewald & Braband 2017, Komárek & Ludvik 1971, 1972, Peličaric 1970). S. intermedius Chodat y S. nanus Chodat, morfológicamente muy similares pudieron ser diferenciadas por la diferente ultraestructura de la pared celular. Como dato curioso, S. perforatus Lemmermann y S. tropicus Crow, morfológicamente diferentes, presentan similar ultraestructura de la pared. De modo que la ultraestructura de la pared celular no ha tenido el mismo peso taxonómico.
Todas las especies de Scenedesmus con estructuras en las paredes celulares: espinas, costillas, dentículos, gránulos, etc. han sido ubicadas en el género Desmodesmus (An et al.1999, Hegewald 2000).
Tanto la taxonomía tradicional, estrictamente morfológica, como los avances con las investigaciones ultraestructurales contribuyeron al conocimiento de las especies morfológicas, aunque las aportaciones de la microscopía electrónica de transmisión y de barrido no se consideran morfológicas en el sentido tradicional, sí se basan también en el fenotipo. De manera que podemos resumir que en Scenedesmus s.l. existen morfoespecies: i) con sus caracteres bien definidos, más o menos estables, ii) con caracteres diagnósticos de amplia variabilidad, que solo evaluaciones poblacionales o de la ultraestructura de la pared permiten su separación, iii) caracteres diagnósticos que se superponen con los de otras, por tanto de dudosa existencia y iv) ocurrencia de morfotipos con ubicación taxonómica incierta, en espera de confirmación o de estudios particulares.
Algunos ejemplos de diferentes interpretaciones en especies de Scenedesmus sensu lato.
He querido seleccionar solo algunos ejemplos en morfoespecies del género Scenedesmus s. l. En la actualidad, principalmente por resultados de las investigaciones genético-moleculares, este género ha sido dividido en varios. De manera que por razones puramente didácticas no aplico los nombres actuales en el momento de presentar los ejemplos, ya que dificultaría la comprensión del tema abordado. No obstante, al finalizar cada grupo se ofrece los nombres más actualizados.
S. disciformis-arcuatus-verrucosus.
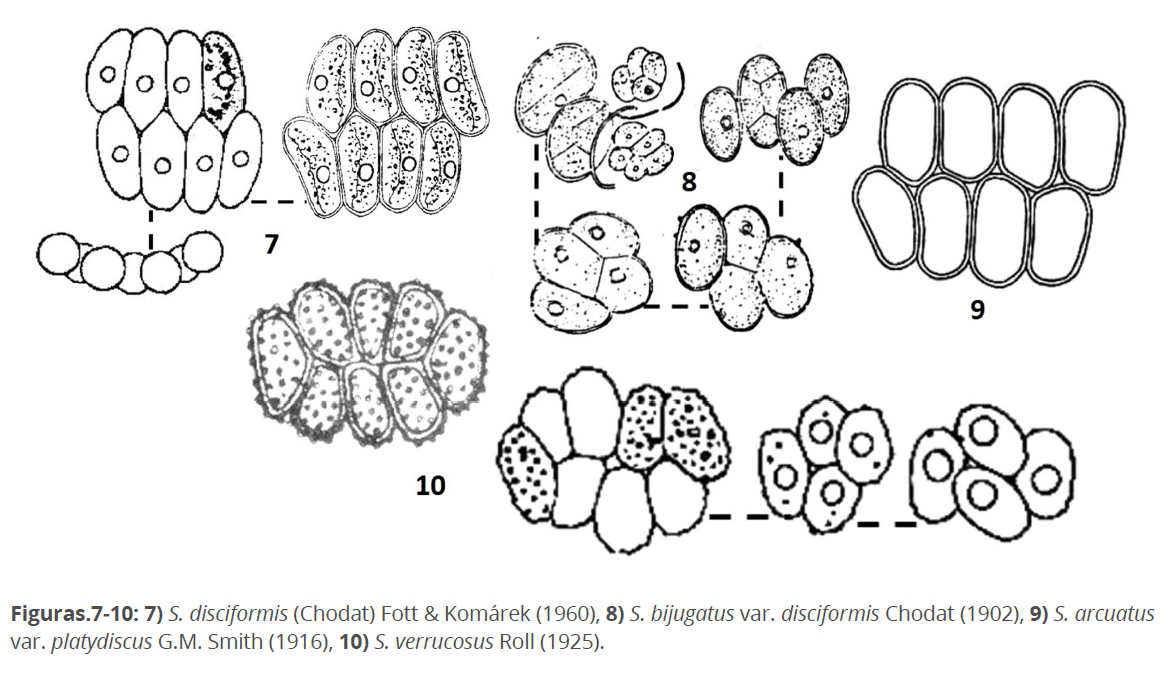
S. disciformis (Chodat) Fott et Komárek 1960 (Fig. 7), tiene como basónimo a S. bijugatus var. disciformis Chodat 1902 (Fig.8) sin embargo, representa a un taxón con cenobios discoidales o con células alternadas pudiendo presentar diminutos dientes. S. disciformis sensu Fott et Komárek posee cenobios con dos hileras de células (con o sin diminutos espacios entre ellas) y carece de dentículos. Este morfotipo existe y ocurre frecuentemente en Cuba y en mi opinión, está relacionado estrechamente con S. arcuatus var. platydiscus G.M. Smith 1916 (Fig. 9), nombrado sin embargo como Comasiella arcuata var. platydisca (Hegewald in Hegewald et al. 2010).
S. bijugatus var. disciformis sensu Chodat, de acuerdo con Hegewald & Schnepf (1978) y Hegewald et al. (1988), es S. verrucosus Roll 1925 (Fig. 10), aunque esta especie no presenta dentículos, solo verrugas que no constituyen estructuras de la pared celular sino deposiciones, que a veces son visibles solo en los márgenes o cubriendo toda la superficie parietal. S. verrucosus se ubica dentro de un género independiente: Verrucodesmus Hegewald et al. 2013.
S. obliquus-dimorphus-acutus.
S. obliquus (Turpin) Kützing 1834, basado en A. obliqua Turpin 1829 (Fig. 11), ha tenido varias interpretaciones. De acuerdo con Hegewald (1982) incluye a S. acutus Meyen 1829 y a S. dimorphus (Turpin) Kützing 1834 (Fig. 12a): Cenobios de 2-4-8 células, alineados, ligera o marcadamente alternados; células más o menos fusiformes hasta cilíndrico-fusiformes, con los extremos atenuados, cónico-puntiagudos con o sin engrosamientos apicales, más o menos rectos o curvos, a veces digitiformes; todas las células iguales, o las marginales lunulares o levemente arqueadas, las internas más o menos rectas; caras celulares de las células externas, rectas o convexas. Esta convexidad sobrepasa o no la línea de los polos. De acuerdo con Komárek & Fott (1983) y Toledo & Comas (1988)
(Fig. 12b-c), presenta células muy variables; pero básicamente oval-delgadas hasta anchamente fusiformes, las externas son más o menos rectas y sus polos son obtusos, no alargados.
S. dimorphus, basado en A. dimorpha Turpin 1828 (Fig. 13), según Komárek & Fott 1983, Toledo & Comas 1988 y varios autores (Fig. 14) está caracterizada por sus células fusiformes, arqueadas, lunulares las externas, si los lados externos de estas células son convexos, esta convexidad no sobrepasa la línea que une a los polos celulares, con polos celulares agudos, cortos o algo pronunciados.
S. acutus, de acuerdo con el iconotipo (Meyen 1829) las células son fusiformes con los polos puntiagudos, las marginales, levemente arqueadas, heteropolares, los márgenes externos de estas células son más o menos rectos (Fig. 15). De acuerdo con diversos autores, la especie presentaba células externas algo curvas hacia fuera del cenobio; pero con sus márgenes externos convexos, convexidad variable, que podía sobrepasar la línea de los polos celulares (Fig. 16a-b).
Estudiando la variabilidad se pueden definir poblaciones que se ajustan a uno de estos tres morfotipos, sin embargo, existen poblaciones intermedias cuya ubicación resulta insegura. La existencia de estas poblaciones intermedias pudiera indicar, siguiendo a la taxonomía tradicional, que se trata de una sola especie con posibilidades de establecer categorías infraespecíficas (Toledo & Comas 1988). Se han incluido dentro de Acutodesmus (Tsarenko in Tsarenko & Petlovanny 2001) y de Tetradesmus (Wynne & Hallan 2015).
S. acuminatus-pectinatus.
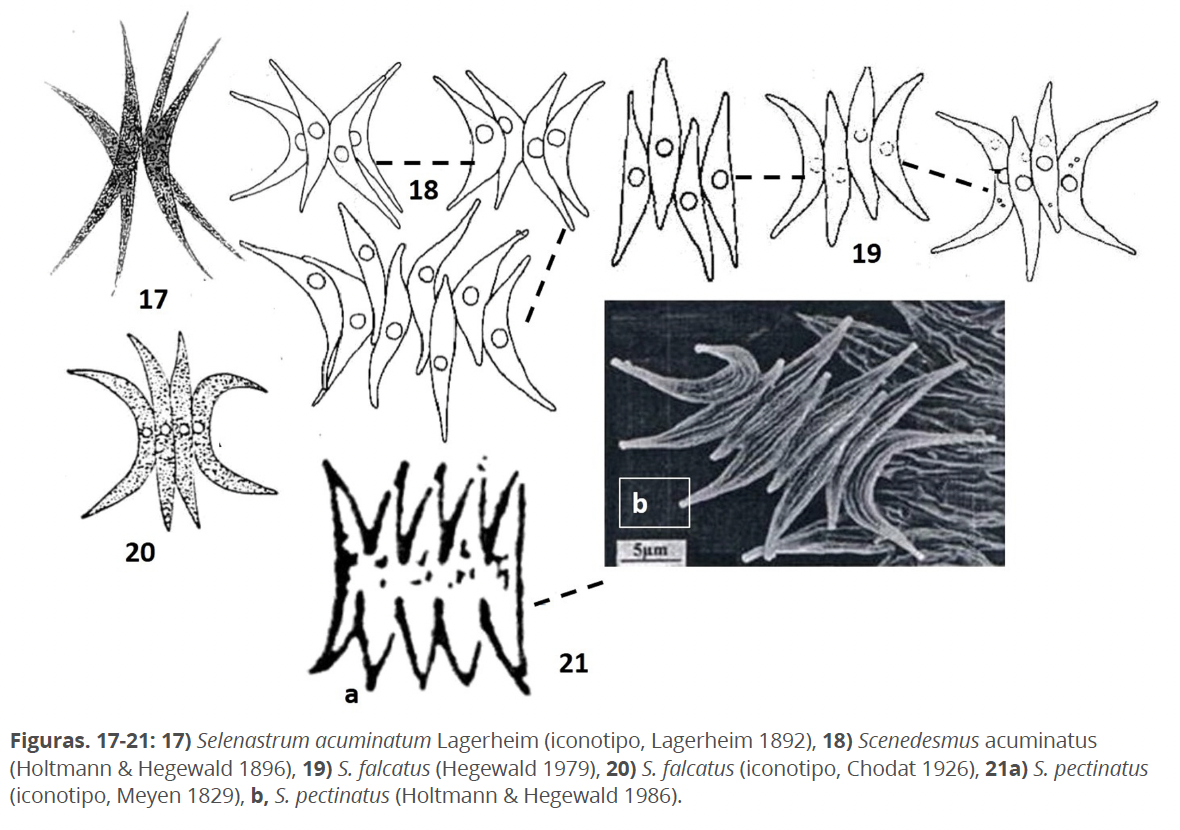
S. acuminatus (Lagerheim) Chodat 1902, basado en Selenastrum acuminatum Lagerheim 1882 (Fig. 17), presenta células fusiformes, delgadas, arqueadas, unidas por sus caras convexas, dispuestas en uno o en diferentes planos, con polos alargados y puntiagudos. Bajo este nombre existían, al menos, dos morfotipos principales, uno con las células delgadas, arqueadas, en diferentes planos, semejante a Selenastrum acuminatum (Fig. 18), al que Hegewald (1979), designa propiamente como Scenedesmus acuminatus, recientemente nombrado Tetradesmus lagerheimii (Wynne & Guiry 2016). El otro morfotipo, con células también arqueadas, delgadas, con polos no tan alargados, formando un cenobio en un mismo plano (Fig. 19), al que Hegewald (1979), aplica primero el nombre de S. falcatus Chodat 1926 (Fig. 20); pero por ilegitimidad del nombre, Holtmann & Hegewald (1986), usan el de S. pectinatus Meyen 1829. El iconotipo de esta especie (Fig. 21a), muestra un taxón, en mi opinión, cercano a S. dimorphus, de hecho, así fue considerado por Komárek & Fott (1983). Como carácter importante, S. pectinatus sensu Holtmann & Hegewald (1986), al microscopio electrónico presenta delicadas costillas en la pared celular (Fig. 21b). No tengo dudas en afirmar que S. pectinatus sensuHoltmann & Hegewald (1986) existe, sin embargo, el nombre aplicado es fruto de una interpretación de S. pectinatus sensu Meyen 1829. A partir de 2013 se incluye en el género Pectinodesmus (Hegewald et al. 2013).
S. bernardii-javanensis-schroeteri.
Bernard (1908), describe para Java (Indonesia) a S. obliquus f. magnus (Fig. 22) que se caracterizaba por sus células fusiformes, delgadas, en cenobios muy alternados (zigzag). G.M. Smith (1916), encuentra en los Estados Unidos una especie con similar formación de cenobios muy alternados, en zigzag, aunque las células eran menos aguzadas y de menores dimensiones (Fig. 23). A pesar de esas diferencias la considera taxonómicamente idéntica a la especie de Bernard (1908), usa el epíteto “bernardii”, ya que “magnus,” a nivel específico existía ya en Scenedesmus (S. magnus Meyen 1829). No designa tipo, ni hace referencia como tal a las figuras de Bernard (1908). Chodat (1926), considera las diferencias morfológicas entre S. bernardii sensu G.M. Smith (1916) y la especie de Bernard (1908), considerándola como una especie diferente: S. javanensis (Fig. 24). Huber-Pestalozzi (1929), describe para Sudáfrica a S. schroeteri, que formaba cenobios en zigzag; pero con células evidentemente mayores y con sus extremos muy alargados (Fig. 25). Comas & Komárek (1984), consideran que S. bernardii, sin tipo designado y diferente al material de Bernard (1908) sería una fuente permanente de error y proponen Scenedesmus pseudobernardii para la especie de G. M. Smith (1916), reteniendo a S. javanensis.
La primera solución fue rechazada por Holtmann & Hegewald (1986), quienes seleccionan como tipo de S. bernardii a la propia diagnosis de G.M. Smith (1916). De acuerdo con Hegewald et al. (2010), S. javanensis pertenece a Pectinodesmus (Hegewald et al. 2010), mientras que S. bernardii ha sido ubicada en Acutodesmus (Hegewald et al. 2013) y en Tetradesmus (Wynne & Hallan 2015).
En mi opinión, en el actual Pectinodesmus javanensis se pueden diferenciar dos tipos morfológicos que coinciden en la formación de cenobios en zigzag; pero se diferencian por la morfología celular y por sus dimensiones: morfotipo “javanensis” (S. javanensis f. javanensis sensu Comas & Komárek 1984) (Fig. 24) y morfotipo “schroeteri” (S. schoeteri f. schroeteri (Huber-Pestalozzi) Comas et Komárek 1984) (Fig. 25).
Una de las más frecuentes interpretaciones ocurre cuando un autor describe una especie cumpliendo con todas las reglas del código vigente. Puede ser considerada dudosa si no se ha encontrado nuevamente después de la descripción original. Ha sucedido más de una vez, que otro autor aplica ese mismo nombre a un morfotipo diferente; pero es adoptado por la mayoría de los autores posteriores. Todo hubiese quedado de esta manera; si no fuese que otro autor halla ambos morfotipos, los estudia y llega a la conclusión de que son buenas especies e independientes, la primera debe mantener el nombre prioritario, la segunda precisa de un nombre nuevo. La selección de este nombre debe ser cuidadosa y consecuente. Tal es el caso de:
S. quadricauda-communis.
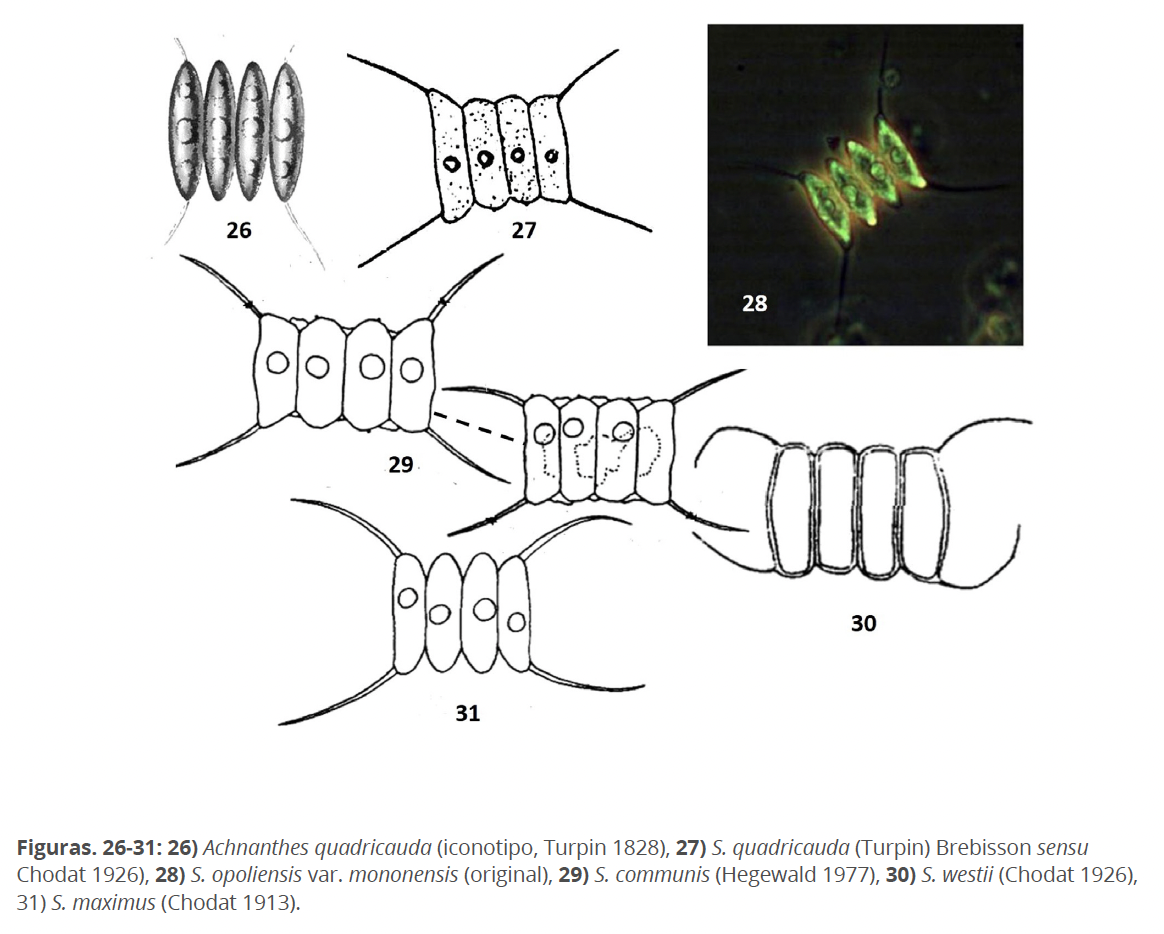
Basado en Achnanthes quadricauda Turpin 1828 (Fig. 26) y formalmente transferido por Brébisson (1835), S. quadricauda (Turpin) Brébisson ha sido una de las especies más comunes y de más amplia distribución, especialmente en los países templados, sin embargo, todos sus hallazgos e identificación se han basado en Chodat (1926), es decir, en una interpretación de este autor (Fig. 27). Hegewald (1977) logra estudiar el material tipo del herbario de Turpin, señaló que aparecían varias especies de Scenedesmus, entre ellas S. opoliensis Richter 1895 (Fig. 28). No halló ningún elemento que coincidiera de manera inequívoca con A. quadricauda. La existencia de S. quadricauda (Turpin) Brébisson en sentido de Chodat (1926), lo motivó a una solución taxonómica, es decir: una nueva especie con nombre nuevo para S. quadricauda sensu Chodat (1926). Para ello, lo más consecuente era seleccionarlo entre los sinónimos anteriores válidamente publicado. Hegewald (1977), descartó a todos los candidatos posibles, hasta llegar a la publicación de S. communis Hegewald (Fig. 29). En mi opinión, S. westii (G.M. Smith) Chodat 1926 (S. quadricauda var. westii G.M. Smith 1916) (Fig. 30) era un candidato posible, sin embargo, lo considera sinónimo de S. maximus (W. et G. S. West) Chodat 1913 (Fig.31), que es en mi opinión diferente.
Según Hegewald (1977), es característica distintiva de S. communis, la membrana entre las células y las rosetas visibles al microscopio fotónico. Ambos caracteres los he observado a veces en poblaciones naturales de S. communis así como también en S. magnus. S. communis fue transferida a Desmodesmus (D. communis (Hegewald) Hegewald 2000). Encuentro gran semejanza entre Achnanthes quadricauda Turpin (Fig.26) y Scenedesmus opoliensis Richter, especialmente con la var. mononensis G.M. Smith 1916 (Fig.28) y soy del criterio de que ambos nombres pudieran pertenecen a una misma especie. Hegewald & Braband (2017), reconocieron esta semejanza. Proponer una solución taxonómica a este problema que además de complicado estaría alejado del objetivo de este trabajo. Menciono además, que Compère & Komárek (1990), propusieron la conservación de S. quadricauda, aceptándose por Greuter et al. (2000), sin embargo, Hegewald & Braband (2017), consideran a esta decisión como no robusta, recomendando su no aplicación (consúltese Hegewald & Braband 2017).
La biología molecular. ¿Solución a las interpretaciones?.
Sin duda el valor científico y los nuevos horizontes que brindan estos estudios avanzados han aclarado y solucionado muchos problemas, entre ellos, las relaciones genético-moleculares entre los grandes grupos y algunos casos a nivel de género y especies. Probablemente el primer estudio de este tipo sobre la familia Scenedesmaceae fue publicado por Paschma & Hegewald (1986)
Sin embargo, algunas soluciones taxonómicas a partir de resultados obtenidos mediante la biología molecular no siempre escapan del subjetivismo. En un inicio, los estudios moleculares se basaron en materiales biológicos identificados previamente siguiendo los criterios tradicionales (morfológicos). Posteriormente estas identificaciones son sustentadas o rectificadas de acuerdo con los resultados genético-moleculares. El clásico ciclo: ¿“qué fue primero, el huevo o la gallina”? Esta coloquial expresión no es tan así.
Primero estuvo la selección de los materiales a estudiar. Al principio, los genetistas moleculares requerían abundante material que fuese genéticamente homogéneo. Fueron entonces a los cultivos depositados en renombradas colecciones. La selección se basó en el nombre de los registros y en las etiquetas correspondientes. Los que hemos tenido la oportunidad de estudiar cultivos de colecciones, sabemos, que no pocas veces, el nombre en las etiquetas no se corresponde con el organismo indicado. El prestigio de esas instituciones estriba fundamentalmente en el estado y conservación de los cultivos; pero por lo general no cuentan con especialistas en cada uno de los grupos taxonómicos, por ejemplo: Hegewald (1989, 1997) en una revisión de la colección UTEX (Universidad de Austin, Texas) de 14 cepas de Scenedesmus rectifica la identificación en 10 de ellas. Los resultados obtenidos mediante los análisis genéticos pueden haber sido científicamente precisos; pero las conclusiones taxonómicas pueden haber sido basadas en materiales incorrectamente identificados. Por experiencia personal, muchas cepas de Coelastrum Nägeli depositadas en colecciones aparecen erróneamente identificadas, lo que ha sido corroborado por Wolf et al. (2003).
Los resultados de estos primeros estudios mostraron que muchos géneros y/o especies presentaban una marcada heterogeneidad, es decir polifiléticos y que los caracteres morfológicos hasta ese momento utilizados por la taxonomía tradicional no definían taxones filogenéticos. Esta última conclusión es correcta. Posteriormente, los equipos de científicos incorporaron taxónomos experimentados que podían ayudar en la selección del material biológico. En esta fase, si los materiales seleccionados se corresponden con los nombres asignados, hay pues garantías en los resultados y en las conclusiones. Existen en la actualidad técnicas de aislamiento que logran obtener cultivos monoclonales que garantizan el material de estudio, no así el nombre aplicado. Debo señalar que, aunque los análisis moleculares sean correctos, si los nombres aplicados son fruto de una interpretación, las conclusiones taxonómico-sistemáticas se apartan de la realidad objetiva. Pudiera abordar otros aspectos sobre los estudios genético-moleculares, concretamente acerca de las conclusiones taxonómico-sistemáticas emanadas de estos. Son aspectos dignos de reflexión; pero me alejaría del tema propuesto para esta contribución.
Para revisiones sobre la taxonomía de los géneros de difícil delimitación, principalmente de aquellos grupos más comunes e importantes en los ecosistemas acuáticos como es el caso de Scenedesmus, es conveniente recordar a los taxónomos la importancia de que las nuevas descripciones se basen en poblaciones representativas, precisa definición de sus caracteres diagnósticos y con una adecuada selección de los tipos nomenclaturales, con una apropiada aplicación del Código Internacional de Nomenclatura para algas, hongos y plantas (ICN). El reconocimiento de especies delimitadas solo por la biología molecular carece de aplicación práctica, no obstante, los estudios moleculares han resultado fundamentales en el entendimiento de las variaciones específicas y tratan de arribar a un concepto de especies dentro de las protistas. Recomiendo las reflexiones de Komárek (2020) sobre cianobacterias que pueden ser aplicadas a todos las microalgas en amplio sentido.
REFERENCIAS
An, S. S., T. Friedl & E. Hegewald, 1999. Phylogenetic relationship of Scenedesmus and Scenedesmus-like coccoid green algae as inferred from ITS-2 rDNA sequence comparisons. Plant Biology 1: 1-11.
Bernard, C. 1908. Protococcacées et desmidiées d’eau douce, récoltées à Java. Batavia, 230 pp.
Bisalputra, T. & T.E. Weier. 1963. The cell wall of Scenedesmus quadricauda. American Journal of Botany 50: 1011-1019.
Brébisson de, A. 1835. Algues des environs de Falaise. Mémoire de la Societé Academie de Falaise, Flaise, 66 pp.
Buchheim, M.A., E. Michalopulus & J. Buchheim. 2001. Phylogeny of Chlorophyceae with special references to the Sphaeropleales: A study of 18S and 26S rDNA data. Journal of Phycology 37: 819-835.
Chodat, R. 1902. Algues vertes de la Suisse. Pleurococcoides et Chroolépiales. - Matériaux Flora Cryptogamie Suisse 1: 373 pp.
Chodat, R. 1913. Monographie d’algues en culture pure. Matériaux Flore Cryptogamie Suisse 4: 1-266.
Chodat, R.1926. Scenedesmus. Étude de génetique, de systématique expérimentale et d’hydrobiologie. Zeitschrift Hydrologie 3: 71-258.
Comas, A. & j. Komárek. 1984. Taxonomy and nomenclature of several species of Scenedesmus (Chlorellales). Algological Studies 35: 135-157.
Compère, P. & j. Komárek. 1990. Proposal to conserve Scenedesmus quadricauda (Turpin) Brébisson with a conserver typ under Art. 69.3 (algae). Taxon 39: 530-531.
Ehrenberg, C.G. 1834. Dritter Beitrag zur Erkenntniss grosser Organisation in der Richtung des kleinsten Raumes. Abhandlungen der Königlichen Akademie der Wissenschaften zu Berlin, Physikalische Klasse 1833: 145-336.
Fott, B. & j. Komárek. 1960. Das Phytoplankton der Teiche im Tschener Schlesien. Preslia 32: 113-141.
Greuter, W., j. Mc Neill, F.R. Barrie, H. Burdet, V. Demoulin, T.S. Filgueiras, D.H. Nicolson, P.C. Silva, J.E. Skog, P. Trehane, N.J. Turland & M.D.L. Hawksworth. 2000. International Code of Botanical Nomenclature (St. Louis code). Regnum Vegetabile 138. Koeltz Scientific Books, Königstein. http://www.bgbm.org/IATP/Nomenclature/Code/SaintLouis/0000St Luistitle.htm
Hegewald, E.1977. Scenedesmus communis Hegew., a new species and its relation to Scenedesmus quadricauda (Turp.) Breb. Algological Studies 19: 142-155.
Hegewald, E. 1979. Vergleichende Beobachtungen and Herbarmaterial und Freilandmaterial von Scenedesmus. Algological Studies 19: 142-155.
Hegewald, E. 1982. Taxonomisch-morphologische Untersuchung von Scenedesmus-Isolaten aus Stammsammlungen. Algological Studies 29: 375-406.
Hegewald, E. 1997. Taxonomy and Phylogeny of Scenedesmus. Algae. The Korean Journal of Phycology 12(4): 235-246.
Hegewald, E. 1989. The Scenedesmus strains of the Culture Collection of the University of Texas at Austin (UTEX). Algological Studies 55: 153-189.
Hegewald, E. 2000. New combinations in the genus Desmodesmus (Chlorophyceae, Scenedesmaceae). Algological Studies 96: 1-18.
Hegewald, E., C. Bock & L. Krienitz. 2013. A phylogenetic study on Scenedesmaceae with the description of a new species of Pectinodesmus and the new genera Verrucodesmus and Chodatodesmus (Chlorophyta, Chlorophyceae). Fottea 14: 149-164.
Hegewald, E. & A. Braband. 2017. A taxonomic revision of Desmodesmus serie Desmodesmus (Sphaeropleales, Scenedesmaceae). Fottea 17: 191-208.
Hegewald, E., N. Jeeji Bai. & M. Hesse. 1975. Taxonomische un floristische Studien an Phytoplanktonalgen aus ungarischen Gewässern. Algological Studies 13: 392-432.
Hegewald, E. & E. Schnepf. 1978. Investigations on the lakes of Perú and their phytoplankton. 3.Three new taxa of Scenedesmus from tropical Perú. Algological Studies 20: 308-317.
Hegewald, E. & P. Silva. 1988. Annotated catalogue of Scenedesmus and nomenclaturally related genera including original descriptions and figures. Bibliotheca Phycologica 80: 1- 587.
Hegewald, E., M. Wolf, A. Keller, T. Friedl, & L. Krienitz. 2010. ITS2 sequence-structure phylogeny in the Scenedesmaceae with special reference to Coelastrum (Chlorophyta, Chlorophyceae), including the new genera Comasiella and Pectinodesmus. Phycologia 49: 325-335.
Holtmann, T. & E. Hegewald 1986. Der Einfluss von Nährlösungen auf die Variabilität von Isolaten der Gattung Scenedesmus. Untergattung Acutodesmus. Archiv für Hydrobiologie. Suppl. 73: 365-380.
Huber-Pestalozzi, G. 1929. Algologische Mitteilungen VI. Archiv für Hydrobiologie 20: 413-426.
Komárek, J. 1978. Taxonomicke Metody. In: Hindák, F. (Ed.). Sladkovodne Riasy, Slovenske Pedagogicke Nakladatelstvo, Bratislava, 197-234 pp.
Komárek, J. 2020. Quo vadis, taxonomy of Cyanobacteria (2019). Fottea 20(1): 104-110.
Komárek, J. & A. Comas. 1982. Taxonomical definition of the genera and several species of Ankistrodesmus and Selenastrum (Chlorococcales). Algological Studies 32: 259-277.
Komárek, J. & B. Fott 1983. Chlorococcales. In: Huber-Pestalozzi (Ed.): Das Phytoplankton des Süsswassers, Systematik und. Biologie”. Teil 7, Stuttgart.
Komárek, J. & J. Ludvik. 1971. Die Zellwandstruktur als taxonomische Merkmal in der Gattung Scenedesmus 1. Die Ultrastrukturelemente. Algological Studies 5: 301-333.
Komárek, J. & J. Ludvik. 1972. Die Zellwandstruktur als taxonomische Merkmal in der Gattung Scenedesmus 2. Übersicht der untersuchten Arten. Algological Studies 6: 11-47.
Kützing, F.T. 1849. Species Algarum, Lipsiae, 922 pp.
Lagerheim, G. 1882. Bidrag till kännedomen om stockholmstraktens Pediastreer, Protococcaceer och Palmellaceer. Öfversigt af Kongl. Vetenskaps-Akademiens Förhandlingar Arg. 39: 47-81.
Lukavský, J. 1991. Motile cells in Scenedesmus obliquus in outdoor mass culture. Archiv Protistenkunde 140: 345-348.
Meyen, F.J.F.1829. Beobachtungen über einige niedere Algenformen. Nova Acta Physico-Medica Academiae Caesareae Leopoldino-Carolinae Naturae 14: 768-778.
Nägeli, C. 1949. Gattungen einzelliger Algen, physiologisch und systematisch bearbeitet. Allgemeine Schweizerisch Gessellschaft für die Gessammte Naturwissenschaften 10(7): 139 pp.
Pelicarič, Š, J. Sulek & J. Ludvik. 1970. Ultrastructure of the cell Wall of Scenedesmus quadricauda (Turp.) Bréb. Strain Greifswald/15. Algological Studies 2/3: 1-6.
Paschma, R. & E. Hegewald. 1986. DNA base composition within the genus Scenedesmus (Chlorophyta). Plant Systematic Evolution 153: 171-180.
Richter, P. 1895. Scenedesmus opoliensis P. Richt, nov. sp. Zeitschrift für angewandte Mikroskopie 1: 3-7.
Roll, J.V. 1925. Les nouvelles espèces des Algues trouvées aux environs de la station biologique du Donetz du Nord. Russkii Arkhiv Protistologii 4: 137-152.
Smith, G. M .1916. A monograph of the algal genus Scenedesmus based upon pure culture studies. Transactions Wisconsin Academy Sciences Arts & Letters 18 (2): 422-530.
Toledo, L. & A. Comas. 1988, Sobre la variabilidad morfológica y la taxonomía de algunas especies de Scenedesmus (Chlorellales). Acta Botánica Cubana 57: 1-32.
Trainor, F.R. 1963. Zoospores in Scenedesmus obliquus. Science 142: 1673-1674.
Tsarenko, P.M. & O.A. Petlovanny. 2001. Droplolneniek Raznoobrazijr vodoroslej Ukrainy. Algologia Suppl.: 1-130.
Turpin, P. J. F. 1828. Apercu organographique sur le nombre deux. Memoire Museum Historia naturell, Paris, 16: 296-344.
Wolf, M., D. Hepperle & L. Krienitz. 2003. On the phylogeny of Planktosphaeria and Schizochlamydella (Radiococcaceae, Chlorophyta). Biologia, Bratislava 58: 759-765.
Wynne, M.J. & J.K. Hallan. 2015. Reinstatement of Tetradesmus G. M. Smith (Sphaeropleales, Chlorophyta). Feddes Repertorium 126: 83-86.
Wynne, M.J. & M.D. Guiry. 2016. Tetradesmus lagerheimii M.J. Wynne & Guiry, nomen novum, a replacement name for Tetradesmus acuminatus (Lagerheim) M.J. Wynne, nom. illeg. (Sphaeropleales, Chlorophyta). Notulae Algarum 12: 1.
Sometido: 13 de noviembre de 2020
Revisado: 12 de febrero (Dra. Thais da Silva y Dra. Rosaluz Tavera)
Corregido: 12 de marzo de 2021
Aceptado: 13 de marzo de 2021